マトリックスメタロプロテアーゼ
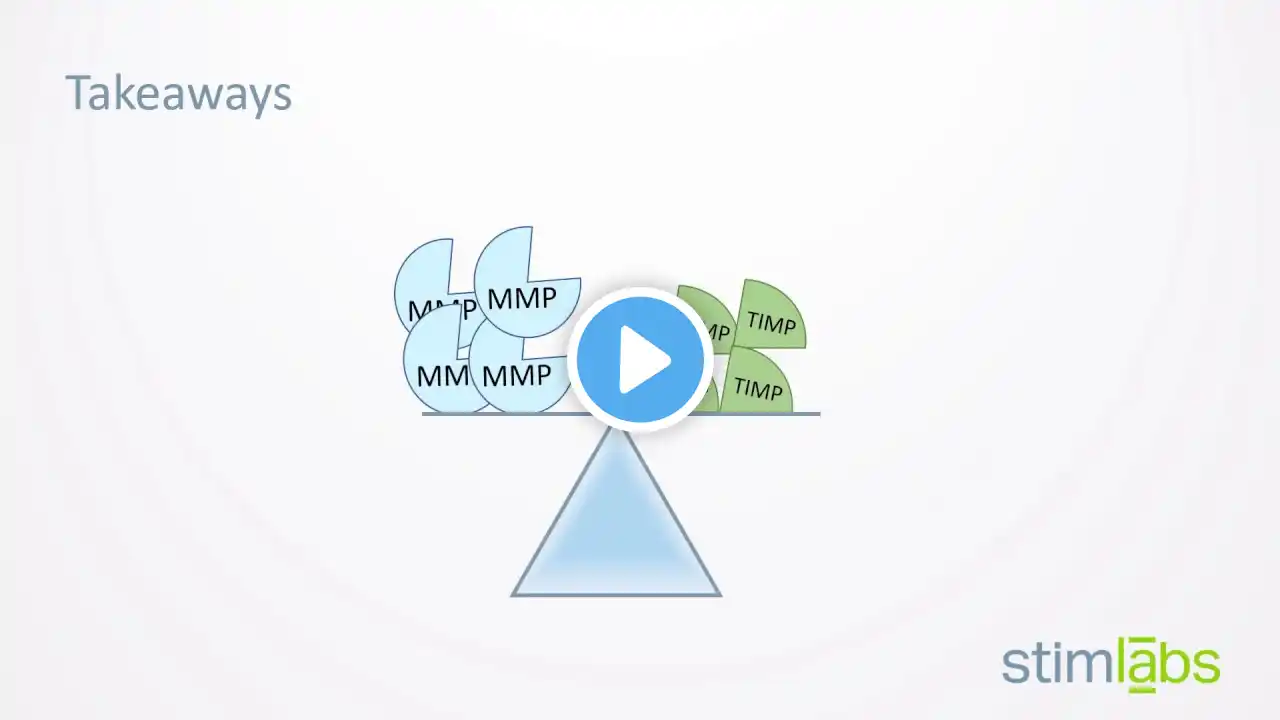
マトリックスメタロプロテアーゼ, by Wikipedia https://ja.wikipedia.org/wiki?curid=1... / CC BY SA 3.0 #ペプチダーゼ #金属タンパク質 マトリックスメタロプロテアーゼ(英:Matrix metalloproteinase、MMP)はメタロプロテアーゼ(活性中心に金属イオンが配座しているタンパク質分解酵素の総称)の一群でありMMPの活性中心には亜鉛イオン(Zn2+)やカルシウムイオン(Ca2+)が含まれる。 コラーゲンやプロテオグリカン、エラスチンなどから成る細胞外マトリックスの分解をはじめとし、細胞表面に発現するタンパク質の分解、生理活性物質のプロセシングなどその作用は多岐にわたる。 1962年にジェロム・グロスとチャールズ・ラピエールによりオタマジャクシの変態において尾が吸収される過程に関与する酵素として発見され、1968年にはヒトの皮膚に存在することが示された。 MMPファミリーに属する酵素は分泌型と膜結合型の二種類に分類される。 分泌型MMPは産生後、分泌細胞から離れたところにおいても働くが、膜結合型は細胞表面に発現しているので活動範囲は狭い。 MT-MMP(MMP14)のカルボキシル基(-COOH)側末端に緑色蛍光タンパク質(GFP)を付け可視化。 MMPファミリーに属する分子は酵素活性ドメインやヘモペキシン様ドメインなどのそれぞれに共通したドメイン構造を有する。 MMPはそれぞれ若干ドメイン構造が異なるが、大まかにはアミノ基(-NH2)末端側からシグナルペプチド、プロペプチド、酵素活性ドメイン、ヒンジ領域、ヘモペキシン様ドメインと続く構造をとる。 なお、シグナルペプチド配列は翻訳後に除去される。 前述したとおりMMPは酵素前駆体として産生され、アミノ基側末端のプロペプチド部位が除去されることにより酵素活性を示す。 プロペプチド部位にはシステインスイッチと呼ばれるアミノ酸配列(PRCGVP)が存在し、よく保存されたシステイン残基が含まれる(MMP-23は例外)。 このシステイン残基のチオール基(-SH)が酵素活性部位の亜鉛と静電気的な相互作用をすることにより基質との結合及び切断を防ぎ、前駆体としての構造を保っている。 酵素活性ドメインにはよく保存された3つのヒスチジン残基があり、酵素活性の発現において重要である亜鉛との結合に関与している。 酵素活性ドメインはヘモペキシン様ドメインと75アミノ酸残基以下のリンカードメインを介してつながっている。 このリンカー部位をヒンジ領域と呼び、柔軟な構造をしている。 ヘモペキシン様ドメイン(英:Hemopexin-like Domain、HLD)はカルボキシル基側末端に存在し、血清タンパク質であるヘモペキシンと相同性がある。 HLDは基質の認識に関与する配列である。 HLDには4枚羽のβプロペラ構造が存在し、タンパク質間の相互作用に関与している。 MMP-7、MMP-23、MMP-26及び植物・線虫のMMPではHLDを欠失している。 MMPの相互活性化経路 MMPは酵素前駆体(チモーゲン)として産生され、プロペプチド部分の酵素的な切断を受けることにより活性化される。 また、RXKRもしくはRRKR配列を持つMMP(膜結合型MMPは全てこの中に含まれる)はゴルジ体に存在するフューリン(英:Furin)と呼ばれる酵素による切断を受け、活性化される。 MMP-1、3、7、9、11、12及び13は誘導型の酵素であり、炎症性刺激により活性化される。 MMPは単に細胞外マトリックスを分解するにとどまらず、サイトカインなどの生理活性ペプチドの活性化など様々な生理現象に関与している。 MMPの基質には様々なものがあり、MMPは骨リモデリングや創傷治癒など生理現象のみならず、炎症や癌の進行などの病的過程にも関与している。 内因性のMMP阻害因子としてTIMP(Tissue Inhibitor of Metalloproteinase)が知られている。 TIMPはMMPと複合体を形成することによりMMPの活性を抑える働きをもっており、過剰なMMPの作用発現による組織破壊を防いでいる。 TIMPファミリーにはTIMP1-4の4種類の分子が属している。 一方、TIMPが過剰に作用することによりMMPによる組織コラーゲンの分解が抑制されて沈着し、繊維化が生じる原因となる。 このようにMMPとTIMPのバランスの維持が重要である。 また、TIMPはMMPだけでなくADAMsファミリーも阻害する。